
写在前面
这个内容在2016年就开始做了,没想到到今天结果才出来。我们使用了不同于五年前的分析技术进行特征微生物的挑选和不同的代谢物验证,终于解析出来了病原菌驱动的黄瓜根际抑病土壤的形成过程。
在这项工作的需求下,为了更好的呈现时间序列和空间序列的微生物组数据分析,我们组将代码使用tidyverse风格重新编写(ps,这里简单介绍一下:tidyverse形式是目前最先进,最为优雅,并且最适合于数据探索的数据结构和形式。目前只存在于R语言中。);此外,当时正值周集中老师发表的NCC的结果分析了微生物团体在网络中的变化,ggClsusterNet的成熟和model_maptree2算法完成。这些共同支持了后续的分析。最终,我们小组使用了网络模块化分析和网络模块相似度检验,结合ggClsusterNet可视化等分析,发现了特征模块在时间梯度下,在抑病土壤形成过程中的变化情况。在没有做任何OTU去除的基础上,可以清楚的看到目标微生物群体的全部变化。
01 摘要
背景:抑病型土壤模型预测植物与其病原体的对抗会导致有益微生物的补充和积累。但哪些有益微生物被富集,以及如何实现对作物病害的抑制,还需要更深入的研究。
方法:我们通过在分根装置中连续种植八季黄瓜并在其一侧接种尖孢镰刀菌来调节土壤。利用宏基因组与非靶向代谢组学等技术挖掘抑病土壤形成中的关键微生物与关键代谢物,再通过多项指标的测定,共同解析两者在抑病土壤形成过程中的主要机制。
结果:病原体感染后,随着根中活性氧(ROS,主要是OH·)的增加以及芽孢杆菌和鞘氨醇单胞菌的积累,疾病发病率逐渐降低。宏基因组测序揭示这些关键微生物可以通过富集双组分系统、细菌分泌系统和鞭毛组装等通路,诱导根部维持高ROS水平,从而保护黄瓜免受病原体感染。非靶向代谢组学分析结合体外应用分析表明,苏糖酸和赖氨酸是招募芽孢杆菌和鞘氨醇单胞菌的关键。
结论:我们的研究破译了一个“cry for help”案例,在这个案例中,黄瓜释放特定的化合物来富集有益的微生物,从而维持宿主的高ROS水平,以防止病原体的入侵。更重要的是,这可能是支撑抑病土壤形成的基本机制之一。
02 前言
在当代集约化全球农业的背景下,维持土壤健康是维系农业生产力的重大挑战。土壤微生物是保持土壤健康的关键,其典型效益是抑制疾病。据报道,抑病型土壤具有抵抗病害发生的能力,即使在病原体存在的情况下也很少或根本没有病害发生。通常情况下,抑病土壤可大致分为一般型抑病土壤和专一型抑病土壤。近几十年来,对专一型抑病型土壤的研究较多,例如参与防治马铃薯赤霉病、小麦全蚀病、番茄青枯病和甜菜青枯病等。这些研究都突出了土壤微生物在病害防治中的重要作用。例如,假单胞菌属通过产生2,4 -二乙酰间苯三酚、吩嗪、吡咯腈、氰酸等生物活性物质介导土壤管理措施。芽孢杆菌属是另一类研究较多的微生物,通过分泌脂肽、表面活性剂和小分子挥发物(VOCs)赋予土壤管理措施。专一型抑病土壤通常是由易感宿主在连续培养过程中爆发的疾病引起的。因此,病原菌、寄主植物和土壤微生物之间的三方相互作用对于特定病害抑制的发生和持续是必要的。根系分泌物作为植物-微生物组通讯的关键媒介,能够响应病原菌并招募有益微生物的帮助。根系分泌物对根际微生物组的正调控作用已得到公认,但病原菌、有益微生物和根系分泌物在专一型抑病土壤形成过程中的确切关系目前尚不清楚。作为一种快速的防御反应,ROS介导的植物-微生物互作受到越来越多的关注。例如,ROS参与根瘤菌-豆科植物共生体系,枯草芽孢杆菌一般会增强宿主植物细胞ROS的产生等。最近发现B. velezensis SQR9 通过上调双组分调控系统诱导植物体内ROS爆发并耐受氧化胁迫。然而,很少有研究评估氧化爆发在土壤微生物组贡献的土壤抑病性中的作用。我们通过在分根系统中连续培养受病原菌胁迫的黄瓜,排除了病原菌对土壤微生物组的直接影响,从而产生了一种抑病土壤。通过宏基因组测序和非靶向代谢组学研究,探索潜在有益菌和根系分泌物在抑病土壤形成中的作用。为了了解病原菌压力下抑病性土壤的形成,我们旨在探究( 1 )病原菌对植物的持续影响富集了哪些微生物? ( 2 )植物分泌物在富集这些微生物中的作用? ( 3 )富集的微生物通过什么机制保护宿主植物抵御病害?
03 结果
1. 病原菌作用下黄瓜抑病土壤的微生物鉴定与特性分析
为了形成专一型的枯萎病抑病土壤,我们在分根装置中进行了黄瓜连作并在一侧根部每季接种尖孢镰刀菌的试验。在8季中,未接种病原菌的土壤(C8)中生长的黄瓜表现出对枯萎病的易感性,发病率为67.59 %,而接种病原菌的土壤中生长的黄瓜发病率从第1季(F1)的51.85 %下降到第8季(F8)的11.11 %(Fig 1A)。Pearson线性回归分析表明在无病原菌处理中连作代数与枯萎病发病率呈显著正相关(P < 0.001 ; R = 0.94),而在病原菌处理中呈负相关(P < 0.001 ; R = -0.88) (Fig 1A )。为了表征抑病性发展过程中微生物组的组装动态,对添加和未添加病原菌的第1、5和8季土壤样品进行DNA提取和16S rRNA测序。α多样性在对照组中随着处理时间的延长而显著降低,而在病原菌处理的样品中则相反(Fig 1B)。然后,我们探究了对照和处理土壤中PCoAs第一轴捕获的β多样性的纵向变化趋势。PCo1解释了对照和病原菌处理样品微生物组分离总变异的26.69 %,进一步揭示了有无病原菌处理下土壤微生物群落组成在第1代和第8代之间的差异显著性(Fig 1C)。为了鉴定随时间变化的受病原菌驯化影响的微生物分类群,对添加和未添加病原菌的第1、5、8季土壤样品中单个ASV的丰度用负二项模型拟合,并与对照处理的Wald检验进行两两比较。在至少一个比较中,共有1042个ASVs受到病原体的影响(Fig. 1D , PFDR < 0.05)。为了进一步确定抑病土壤形成过程中差异丰富的ASVs的一致性模式,对所有组之间计算的log2倍变化进行了层次聚类,并区分了10个聚类,以显示不同组之间的不同趋势(Fig. 1D)。对于每个聚类,发现每季纵向趋势的平均log2倍变化中”Cluster 2 “趋势与枯萎病发病率显著相关(R = 0.902; P < 0.001) (Fig. 1E)。” Cluster 2 “由48个属的414个ASVs组成,它们的总相对丰度在F1至F8中随着抑病土壤的形成而增加,而在C1至C8土壤中则相反。对细菌相对丰度和发病率进行Beta回归,发现7个属(芽孢杆菌属、土壤杆菌属、假单胞菌属、鞘氨醇单胞菌属、假黄单胞菌)与发病率显著相关。
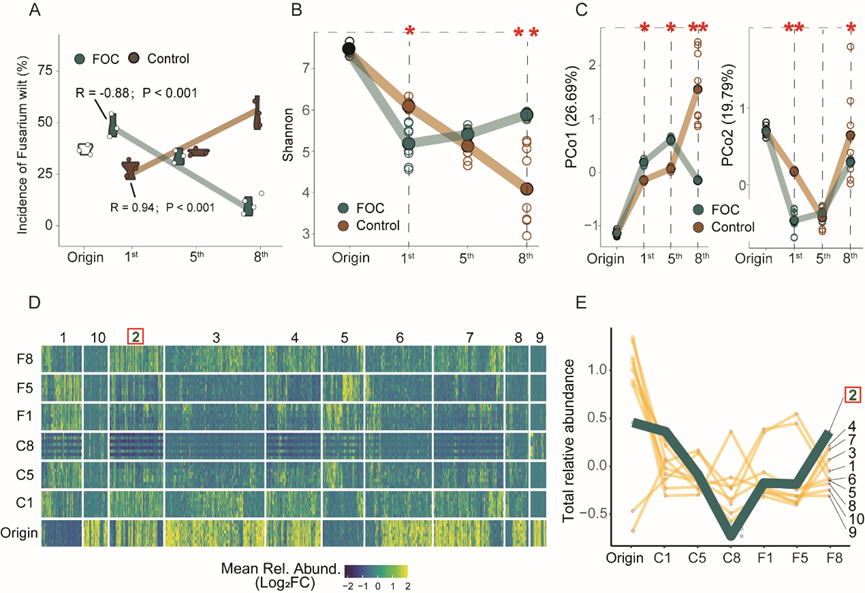
Fig 1 黄瓜连作8季的发病率和微生物群落贡献
2. 关键微生物的分离及其生物防治机制的研究
网络分析发现,F8样品中hub ASVs主要属于微小杆菌属(1个ASV )、Blastomona(1个ASV )、Bacillus(3个ASV )和鞘氨醇单胞菌属(1个ASV )。这与芽胞杆菌和鞘氨醇单胞菌属在形成” Cluster 2 “的7个属中的存在显著相关,表明芽胞杆菌和鞘氨醇单胞菌属在抑病土壤中起关键作用。基于网络的模块分析表明,假单胞菌属、鞘氨醇单胞菌属、芽孢杆菌属和红游动菌属是F8样品最大模块中的主要成员。同时含有芽孢杆菌和鞘氨醇单胞菌的模块在C1样品中比在F8样品中更丰富(Fig 2A),并且根据模块相似性分析,许多C1模块与F8中的模块相似。这些模块的频率从C1到C8逐渐降低,从F1到F8逐渐增加。因此,我们将芽孢杆菌和鞘氨醇单胞菌定义为在土壤抑制作用发展中具有潜在重要功能作用的关键微生物。为了证明因果关系,更好地了解关键微生物如何协助植物防止病原体感染,我们从F8土壤样品中分离了102株细菌,获得了6株芽孢杆菌属和5株鞘氨醇单胞菌属菌株。应用这些菌株的人工复合菌系降低了枯萎病发病率,其中两种微生物组的混合物的发病率最低(Sym_BS : 17.93 %) (Fig 2B)。为了解析这些微生物的生防机制,首先我们评估后发现它们对尖孢镰刀菌不具有直接拮抗特性,再通过测定防御相关的植物激素发现在对照组和微生物处理组中,诱导性系统抗性(ISR)和系统获得性抗性(SAR)之间没有显著差异。然而,接种Sym_BS的黄瓜根系比对照组根系中表现出更高的活性氧水平(Fig 2C)。通过测试ROS的主要组成成分(OH·、O2-和H2O2),发现OH·是ROS增加的主要原因(Fig 2D)。为了进一步评估维持宿主根部高水平ROS的微生物是否负责抑制土壤中的病原菌,将黄瓜幼苗播种到有无病原菌处理的第1、5和8季后收集的土壤中。结果表明,当黄瓜生长在病原菌(F1 ~ F8)持续侵染的土壤中时,OH·水平逐渐升高,而在对照土壤(从C1到C8)中则无显著差异(Fig 2E)。线性回归分析表明,OH·含量与关键微生物丰度呈显著正相关(Fig 2F : R = 0.78 , P < 0.001),与枯萎病发病率呈负相关(Fig 2G : R = -0.81 ; P < 0.001)。这些结果表明芽孢杆菌和鞘氨醇单胞菌提高了黄瓜根部OH·的含量,从而降低了发病率。
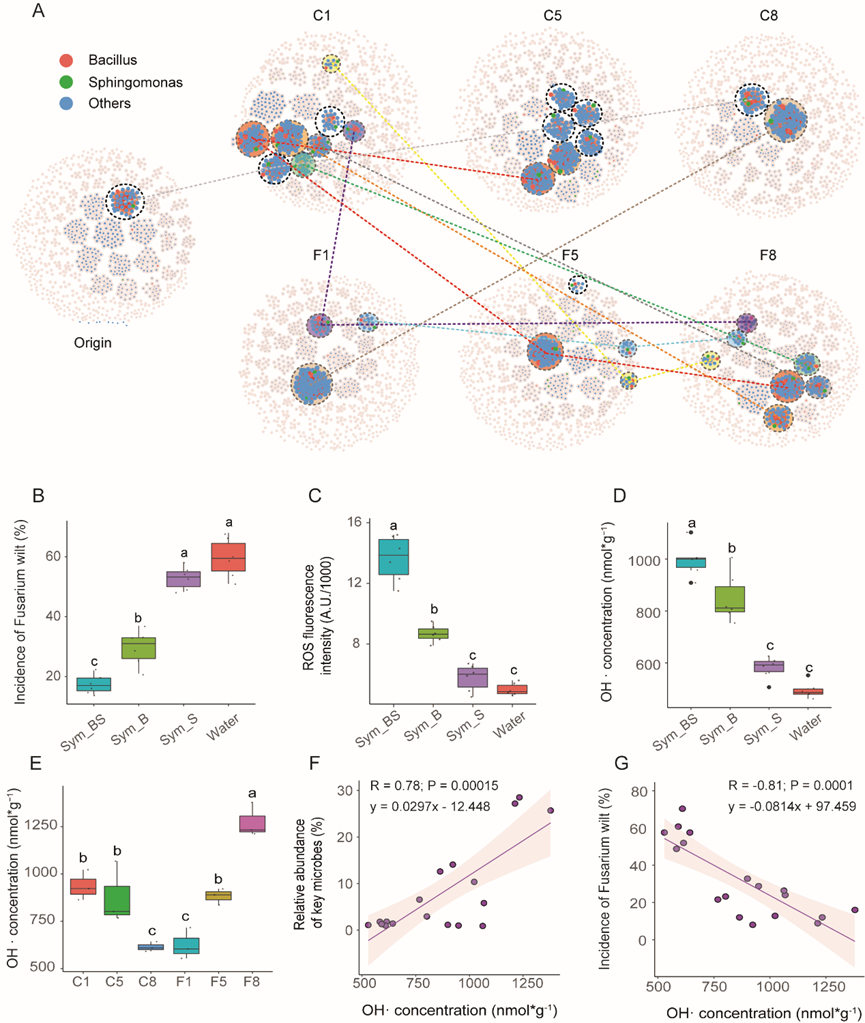
Fig 2 关键微生物的研究及其功能展示
3. 尖孢镰刀菌感染可刺激苏糖酸和赖氨酸募集芽孢杆菌和鞘氨醇单胞菌
为了确定芽孢杆菌和鞘氨醇单胞菌属是如何在病原菌的持续处理下富集的,我们比较了接种和不接种病原菌的黄瓜根系分泌物。通过Kruskal-Wallis非参数检验和倍数差异分析(log2FC > 2),发现其中30个代谢物在两组间具有显著差异 (P < 0.05) (Fig 3A)。其中只有苏糖酸和赖氨酸促进了混合菌株的体外生长(Fig 3B)。使用苏糖酸和赖氨酸预处理(培育)土壤,其中微生物组降低了枯萎病菌的发病率(Fig 3C),在添加代谢物的土壤条件下生长的黄瓜根系的OH·含量也高于对照组(Fig 3D)。本试验中,OH·含量与枯萎病发病率之间也存在显著的负相关关系(Fig 3E , R = -0.98 ; P < 0.001)。此外,施用苏糖酸和赖氨酸显著提高了芽孢杆菌和鞘氨醇单胞菌属的相对丰度(Fig 3F)。
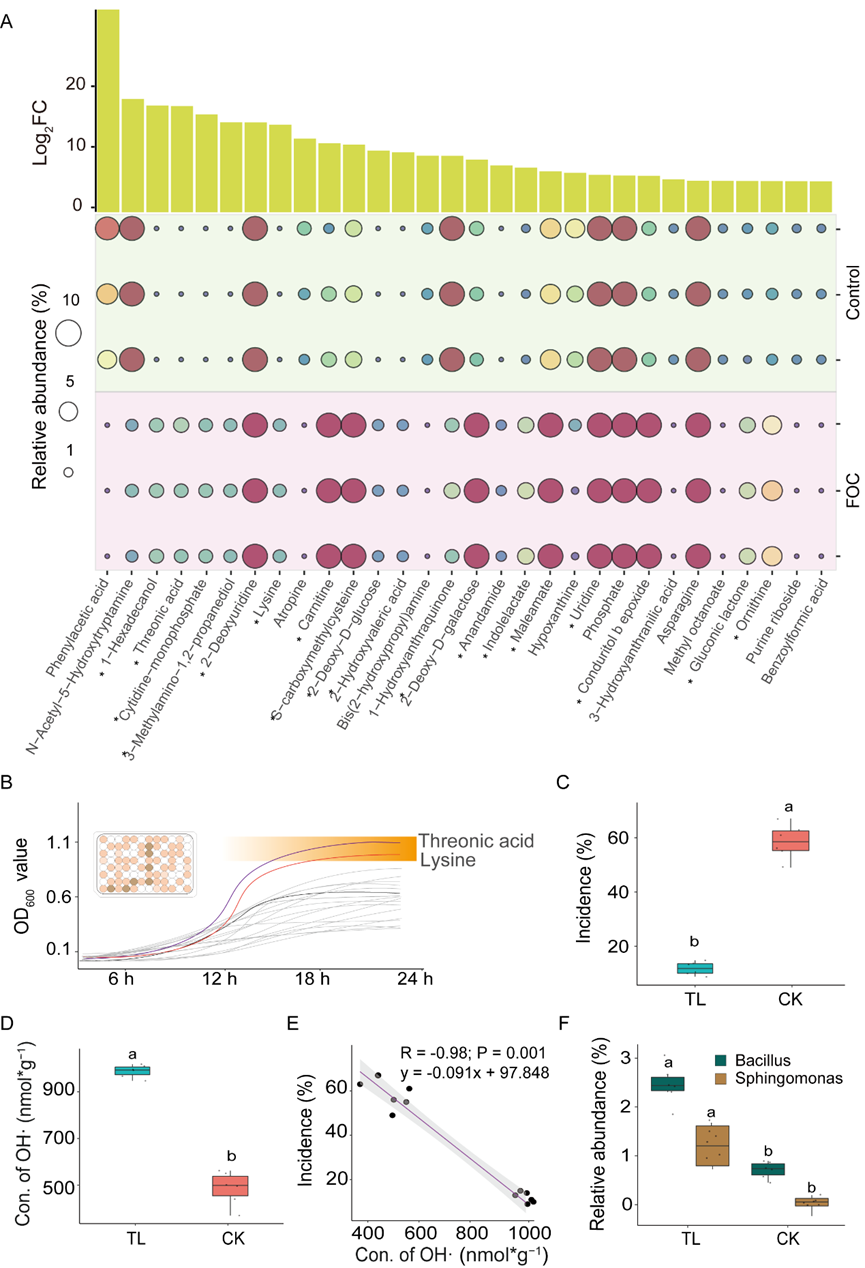
Fig 3 关键代谢产物研究及其对关键微生物的影响
4. 黄瓜抑病土壤的功能分析及芽孢杆菌和鞘氨醇单胞菌的功能分析
为了揭示该抗病土壤中芽孢杆菌属和鞘氨醇单胞菌属等生物群落的潜在功能特征,比较了F8和C8土壤样品的宏基因组图谱。基于Bray-Curtis距离的PCoA显示C8和F8的微生物特征之间存在显著差异(MRPP , P = 0.001 , delta = 0.091) (Fig 4A)。Limma方法的变异分析表明,F8样品中富集的微生物群包括乳酸杆菌、葡萄球菌、芽孢杆菌、鞘氨醇单胞菌和假单胞菌(Fig 4B)。基因集变异分析(GSVA)揭示了一些功能,如双组分系统(ko02020)、细菌分泌系统(ko03070)、鞭毛组装(ko02040)、细菌趋化性(ko02030)和蛋白质输出(ko03060),以及其他途径(赖氨酸降解、脂肪酸代谢、丙氨酸、天冬氨酸和谷氨酸代谢等)在F8土壤样品中富集。然后,提取了芽孢杆菌和鞘氨醇单胞菌属的基因,发现它们对双组分系统、鞭毛组装、细菌趋化性和赖氨酸降解的功能变异具有重要贡献。同时,大部分直接参与鞭毛组装的基因在F8中富集(Fig 4C)。此外,间接参与鞭毛组装的基因也在F8中富集,包括细菌趋化通路(MCP ; che A ; cheY)和双组分系统(flr C) (Fig 4C)。

Fig 4 八季土壤微生物群落在病原体处理和对照之间的功能特征
5.结论
本研究以黄瓜为模型,探究了抑病型土壤的形成机制。本研究发现,芽胞杆菌和鞘氨醇单胞菌属在此过程中富集,并诱导黄瓜根部维持高水平的ROS,尤其是OH·,对黄瓜枯萎病具有保护作用。进一步分析表明,关键微生物可以激活包括双组分调控系统、细菌分泌系统和鞭毛合成与组装基因在内的通路。黄瓜根系分泌物组分苏糖酸和赖氨酸可能是参与招募关键微生物响应病原菌入侵的主要因素,因为这些微生物具有赖氨酸降解的代谢途径。施用苏糖酸和赖氨酸后,黄瓜枯萎病发病率降低至20%,根际芽孢杆菌显著富集,黄瓜根部ROS(OH·)水平较高。我们的研究为今后阐明抑病型土壤形成的机制提供了重要的理论依据,从而为土传病害的防治提供了有效的途径。
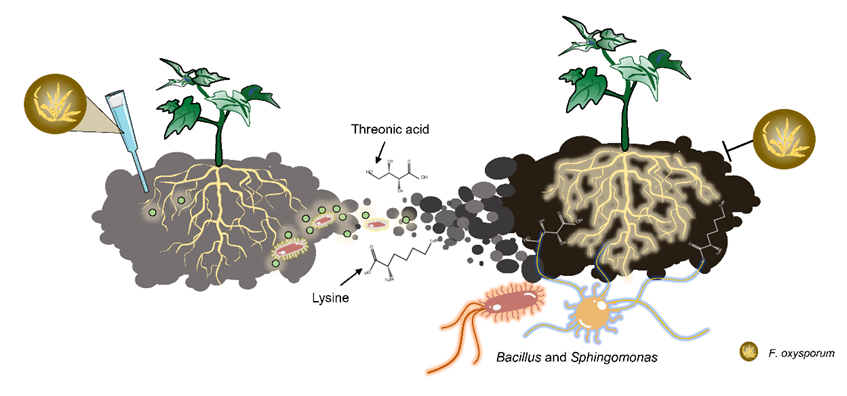
Fig 5 寄主植物根系中活性氧爆发的示意模型
原文链接:https://mp.weixin.qq.com/s/XgJ4mcZfExUHVISr0y3i-Q